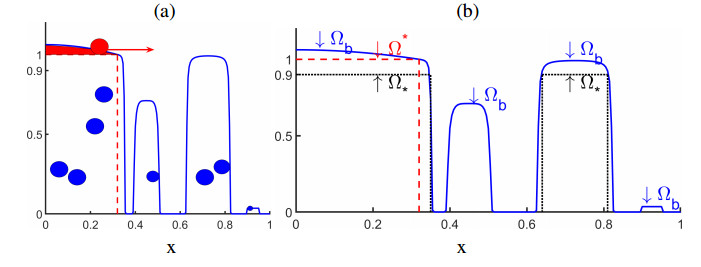
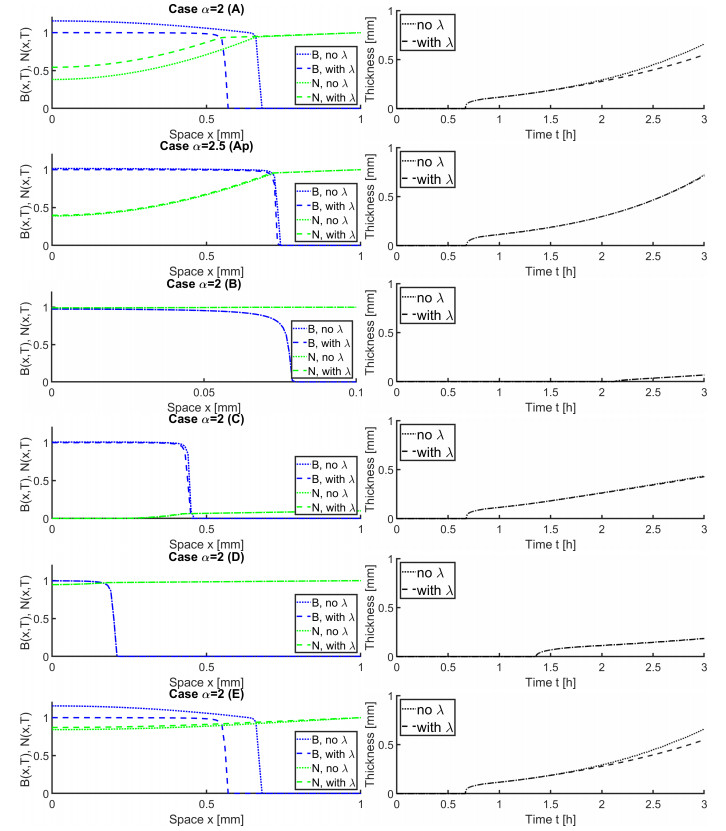
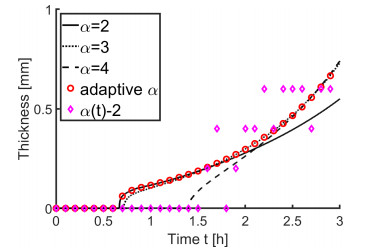
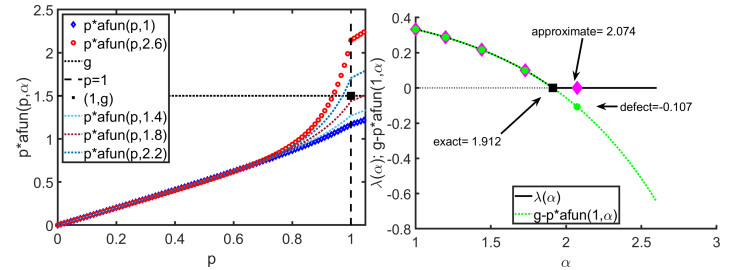
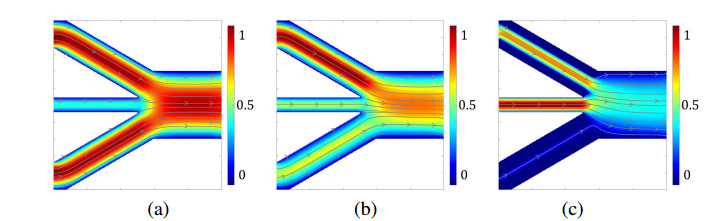
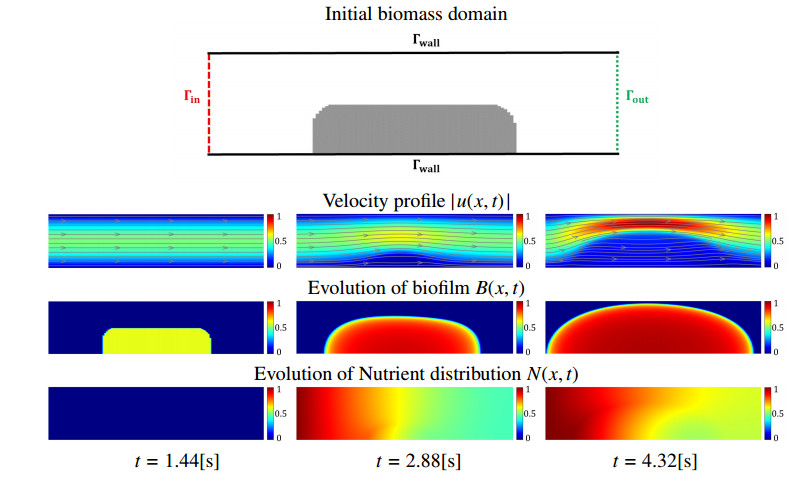
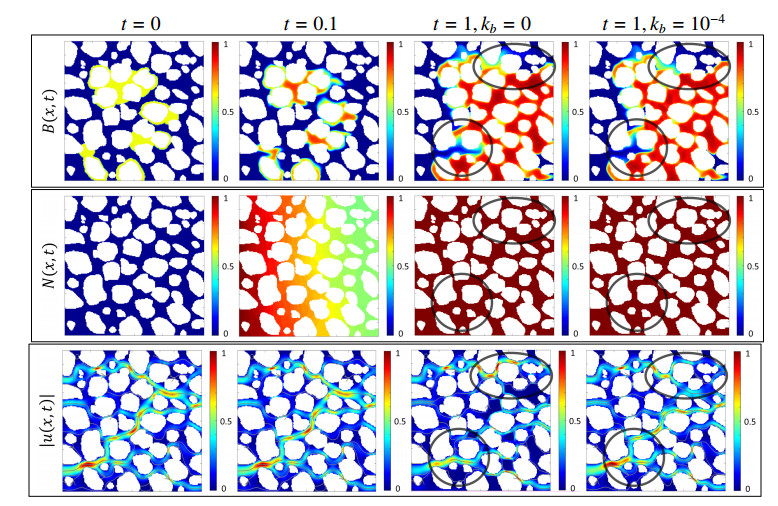
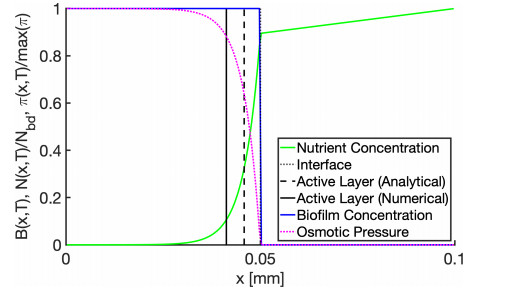
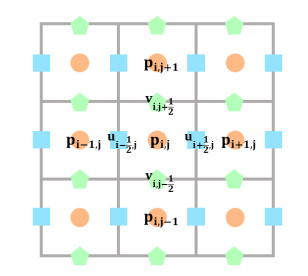
In this paper we describe a coupled model for flow and microbial growth as well as nutrient utilization. These processes occur within and outside the biofilm phase formed by the microbes. The primary challenge is to address the volume constraint of maximum cell density but also to allow some microbial presence outside the contiguous biofilm phase. Our model derives from the continuum analogues of the mechanism of cell shoving introduced in discrete biomass models, and in particular from the models exploiting singular diffusivity as well as from models of variational inequality type which impose explicit constraints. We blend these approaches and propose a new idea to adapt the magnitude of the diffusivity automatically so as to ensure the volume constraint without affecting the reactions; this construction can be implemented in many variants without deteriorating the overall efficiency. The second challenge is to account for the flow and transport in the bulk fluid phase adjacent to the biofilm phase. We use the Brinkman flow model with a spatially variable permeability depending on biomass amount. The fluid flow allows some advection of the nutrient within the biofilm phase as well as for the flow even when the pores are close to being plugged up. Our entire model is monolithic and computationally robust even in complex pore-scale geometries, and extends to multiple species. We provide illustrations of our model and of related approaches. The results of the model can be easily post—processed to provide Darcy scale properties of the porous medium, e.g., one can predict how the permeability changes depending on the biomass growth in many realistic scenarios.
Citation: Choah Shin, Azhar Alhammali, Lisa Bigler, Naren Vohra, Malgorzata Peszynska. Coupled flow and biomass-nutrient growth at pore-scale with permeable biofilm, adaptive singularity and multiple species[J]. Mathematical Biosciences and Engineering, 2021, 18(3): 2097-2149. doi: 10.3934/mbe.2021108
In this paper we describe a coupled model for flow and microbial growth as well as nutrient utilization. These processes occur within and outside the biofilm phase formed by the microbes. The primary challenge is to address the volume constraint of maximum cell density but also to allow some microbial presence outside the contiguous biofilm phase. Our model derives from the continuum analogues of the mechanism of cell shoving introduced in discrete biomass models, and in particular from the models exploiting singular diffusivity as well as from models of variational inequality type which impose explicit constraints. We blend these approaches and propose a new idea to adapt the magnitude of the diffusivity automatically so as to ensure the volume constraint without affecting the reactions; this construction can be implemented in many variants without deteriorating the overall efficiency. The second challenge is to account for the flow and transport in the bulk fluid phase adjacent to the biofilm phase. We use the Brinkman flow model with a spatially variable permeability depending on biomass amount. The fluid flow allows some advection of the nutrient within the biofilm phase as well as for the flow even when the pores are close to being plugged up. Our entire model is monolithic and computationally robust even in complex pore-scale geometries, and extends to multiple species. We provide illustrations of our model and of related approaches. The results of the model can be easily post—processed to provide Darcy scale properties of the porous medium, e.g., one can predict how the permeability changes depending on the biomass growth in many realistic scenarios.

| [1] |
R. Sacco, P. Causin, C. Lelli, M. T. Raimondi, A poroelastic mixture model of mechanobiological processes in biomass growth: theory and application to tissue engineering, Meccanica (Milan), 52 (2017), 3273–3297. doi: 10.1007/s11012-017-0638-9

|
| [2] |
N. G. Cogan, J. P. Keener, The role of the biofilm matrix in structural development, Math. Med. Biol.: J. IMA, 21 (2004), 147–166. doi: 10.1093/imammb/21.2.147
|
| [3] |
T. Zhang, N. G. Cogan, Q. Wang, Phase-field models for biofilms. ⅰ. theory and one-dimensional simulations, SIAM J. Appl. Math., 69 (2008), 641–669. doi: 10.1137/070691966
|
| [4] |
T. Zhang, I. Klapper, Mathematical model of biofilm induced calcite precipitation, Water Sci. Technol., 61 (2010), 2957–2964. doi: 10.2166/wst.2010.064
|
| [5] |
T. Zhang, I. Klapper, Mathematical model of the effect of electrodiffusion on biomineralization, Int. J. Non-linear Mechanics, 46 (2011), 657–666. doi: 10.1016/j.ijnonlinmec.2010.12.008
|
| [6] |
B. D. Wood, S. Whitaker, Diffusion and reaction in biofilms, Chem. Eng. Sc., 53 (1998), 397–425. doi: 10.1016/S0009-2509(97)00319-9
|
| [7] | D. H. Limoli, C. J. Jones, D. J. Wozniak, Bacterial extracellular polysaccharides in biofilm formation and function, Microb. Biofilms, (2015), 223–247. |
| [8] | F. S. Colwell, R. W. Smith, F. G. Ferris, A. L. Reysenbach, Y. Fujita, T. L. Tyler, et al., Microbially mediated subsurface calcite precipitation for removal of hazardous divalent cations: microbial activity, molecular biology, and modeling, in ACS Symposium Series, American Chemical Society, (2005), 117–137. |
| [9] |
I. Klappe, J. Dockery, Mathematical description of microbial biofilms, SIAM Rev., 52 (2010), 221–265. doi: 10.1137/080739720
|
| [10] |
F. A. MacLeod, H. M. Lappin-Scott, J. W. Costerton, Plugging of a model rock system by using starved bacteria, Appl. Environ. Microbiol., 54 (1988), 1365–1372. doi: 10.1128/AEM.54.6.1365-1372.1988
|
| [11] |
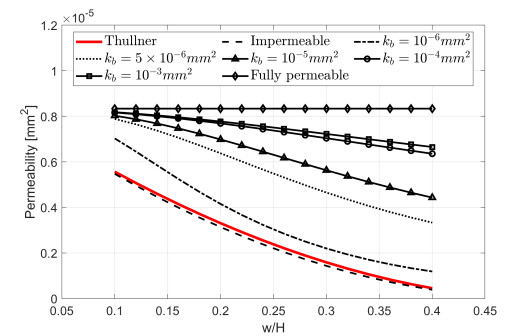
M. Thullner, Comparison of bioclogging effects in saturated porous media within one-and two-dimensional flow systems, Ecol. Eng., 36 (2010), 176–196. doi: 10.1016/j.ecoleng.2008.12.037
|
| [12] |
A. Ebigbo, R. Helmig, A. B. Cunningham, H. Class, R. Gerlach, Modelling biofilm growth in the presence of carbon dioxide and water flow in the subsurface, Adv. Water Resources, 33 (2010), 762–781. doi: 10.1016/j.advwatres.2010.04.004
|
| [13] |
K. Z. Coyte, H. Tabuteau, E. A. Gaffney, K. R. Foster, W. M. Durham, Microbial competition in porous environments can select against rapid biofilm growth, Proc. Natl. Aca. Sci., 114 (2017), E161–E170. doi: 10.1073/pnas.1525228113
|
| [14] |
K. Drescher, Y. Shen, B. L. Bassler, H. A. Stone, Biofilm streamers cause catastrophic disruption of flow with consequences for environmental and medical systems, Proc. Natl. Aca. Sci., 110 (2013), 4345–4350. doi: 10.1073/pnas.1300321110
|
| [15] |
Y. Tang, A. J. Valocchi, C. J. Werth, H. Liu, An improved pore-scale biofilm model and comparison with a microfluidic flow cell experiment, Water Resources Res., 49 (2013), 8370–8382. doi: 10.1002/2013WR013843
|
| [16] | T. L. van Noorden, I. S. Pop, A. Ebigbo, R. Helmig, An upscaled model for biofilm growth in a thin strip, Water Resources Res., 46 (2010), W06505–1/14. |
| [17] |
M. Peszynska, A. Trykozko, G. Iltis, S. Schlueter, D. Wildenschild, Biofilm growth in porous media: Experiments, computational modeling at the porescale, and upscaling, Adv. Water Resources, 95 (2016), 288–301. doi: 10.1016/j.advwatres.2015.07.008
|
| [18] |
I. Klapper, J. Dockery, Finger formation in biofilm layers, SIAM J. Appl. Math., 62 (2002), 853–869. doi: 10.1137/S0036139900371709
|
| [19] |
H. J. Eberl, C. Picioreanu, J. J. Heijnen, M. C. M. van Loosdrecht, A three-dimensional numerical study on the correlation of spatial structure, hydrodynamic conditions, and mass transfer and conversion in biofilms, Chem. Eng. Sci., 55 (2000), 6209–6222. doi: 10.1016/S0009-2509(00)00169-X
|
| [20] |
M. R. Frederick, C. Kuttler, B. A. Hense, H. J. Eberl, A mathematical model of quorum sensing regulated eps production in biofilm communities, Theor. Biol. Med. Model., 8 (2011), 8. doi: 10.1186/1742-4682-8-8
|
| [21] |
D. Landa-Marbán, N. Liu, I. S. Pop, K. Kumar, P. Pettersson, G. Bødtker, et al., A pore-scale model for permeable biofilm: numerical simulations and laboratory experiments, Transp. Porous Media, 127 (2019), 643–660. doi: 10.1007/s11242-018-1218-8
|
| [22] | T. Zhang, N. Cogan, Qi Wang, Phase-field models for biofilms ⅱ. 2-d numerical simulations of biofilm-flow interaction, Commun. Comput. Phys, 4 (2008), 72–101. |
| [23] |
J. B. Xavier, K. R. Foster, Cooperation and conflict in microbial biofilms, Proceedings of the National Academy of Sciences, 104 (2007), 876–881. doi: 10.1073/pnas.0607651104
|
| [24] | A. Alhammali, Numerical Analysis of a System of Parabolic Variational Inequalities with Application to Biofilm Growth, Ph.D thesis, Oregon State University, 2019. |
| [25] |
A. Alhammali, M. Peszynska, Numerical analysis of a parabolic variational inequality system modeling biofilm growth at the porescale, Numer. Methods Partial Differ. Equations, 36 (2020), 941–971. doi: 10.1002/num.22458
|
| [26] |
E. Alpkvist, I. Klapper, A multidimensional multispecies continuum model for heterogeneous biofilm development, Bull. Math. Bbiol., 69 (2007), 765–789. doi: 10.1007/s11538-006-9168-7
|
| [27] |
T. B. Costa, K. Kennedy, M. Peszynska, Hybrid three-scale model for evolving pore-scale geometries, Comput. Geosci., 22 (2018), 925–950. doi: 10.1007/s10596-018-9733-9
|
| [28] | S. J Pirt, A kinetic study of the mode of growth of surface colonies of bacteria and fungi, Microbiology, 47 (1967), 181–197. |
| [29] | M. Efendiev, Evolution Equations Arising in the Modelling of Life Sciences, Springer Basel AG, Basel, 2013. |
| [30] | K. Williamson, P. Mccarty, A model of substrate utilization by bacterial films, Water Pollut. Control Fed., 48 (1976), 9–24. |
| [31] | J. U. Kreft, C. Picioreanu, M. C. M van Loosdrecht, J. W. T. Wimpenny, Individual-based modelling of biofilms, Microbiol. (Soc. General Microbiol.), 147 (2001), 2897–2912. |
| [32] | R. J. LeVeque, Finite Difference Methods for Ordinary and Partial Differential Equations: Steady-state and Time-Dependent Problems, Society for Industrial and Applied Mathematics (SIAM), Philadelphia, PA, 2007. |
| [33] | M. Peszynska, T. Anna, Convergence and stability in upscaling of flow with inertia from porescale to mesoscale, Int. J. Multiscale Comput. Eng., 9 (2011), 215–229. |
| [34] |
M. Peszynska, A. Trykozko, Pore-to-core simulations of flow with large velocities using continuum models and imaging data, Comput. Geosci., 17 (2013), 623–645. doi: 10.1007/s10596-013-9344-4
|
| [35] | T. F. Russell, M. F. Wheeler, Finite element and finite difference methods for continuous flows in porous media, in The Mathematics of Reservoir Simulation (ed. R. E. Ewing), SIAM, Philadelphia, (1983), 35–106. |
| [36] | M. Peszynska, S. Sun, Reactive transport model coupled to multiphase flow models, in Computational Methods in Water Resources (eds. S. M. Hassanizadeh, R. J. Schotting, W. G. Gray and G. F. Pinder), Elsevier, (2002), 923–930. |
| [37] |
N. Nishiyama, T. Yokoyama, Permeability of porous media: role of the critical pore size: critical pore zize-permeability relation, J. Geophysical Res. Solid Earth, 122 (2017), 6955–6971. doi: 10.1002/2016JB013793
|
| [38] |
A. B. Cunningham, W. G. Characklis, F. Abedeen, D. Crawford, Influence of biofilm accumulation on porous media hydrodynamics, Environ. Sci. Technol., 25 (1991), 1305–1311. doi: 10.1021/es00019a013
|
| [39] | S. Bryant, L. Britton, Mechanistic Understanding of Microbial Plugging for Improved Sweep Efficiency, Technical report, The University Of Texas At Austin, 2008. |
| [40] |
S. Schlüter, A. Sheppard, K. Brown, D. Wildenschild, Image processing of multiphase images obtained via x-ray microtomography: a review, Water Resources Res., 50 (2014), 3615–3639. doi: 10.1002/2014WR015256
|
| [41] | H. J. Eberl, L. Demaret, A finite difference scheme for a degenerated diffusion equation arising in microbial ecology, Electron. J. Differ. Equations, 15 (2007), 77–95. |
| [42] |
C. Picioreanu, M. C. M van Loosdrecht, J. J. Heijnen, Mathematical modeling of biofilm structure with a hybrid differential-discrete cellular automaton approach, Biotechnol. Bioeng., 58 (1998), 101–116. doi: 10.1002/(SICI)1097-0290(19980405)58:1<101::AID-BIT11>3.0.CO;2-M
|
| [43] |
C. Picioreanu, M. C. M van Loosdrecht, J. J. Heijnen, A new combined differential‐discrete cellular automaton approach for biofilm modeling: application for growth in gel beads, Biotechnol. Bioeng., 57 (1998), 718–731. doi: 10.1002/(SICI)1097-0290(19980320)57:6<718::AID-BIT9>3.0.CO;2-O
|
| [44] |
E. Alpkvist, C. Picioreanu, M. C. M. van Loosdrecht, A. Heyden, Three-dimensional biofilm model with individual cells and continuum eps matrix, Biotechnol. Bioeng., 94 (2006), 961–979. doi: 10.1002/bit.20917
|
| [45] |
P. G. Jayathilake, S. Jana, S. Rushton, D. Swailes, B. Bridgens, T. Curtis, et al., Extracellular polymeric substance production and aggregated bacteria colonization influence the competition of microbes in biofilms, Front. Microbio., 8 (2017), 1865–1865. doi: 10.3389/fmicb.2017.01865
|
| [46] |
D. Landa-Marbán, G. Bødtker, K. Kumar, I. S. Pop, F. A. Radu, An upscaled model for permeable biofilm in a thin channel and tube, Transp. Porous Media, 132 (2020), 83–112. doi: 10.1007/s11242-020-01381-5
|
| [47] | R. Glowinski, Numerical Methods for Nonlinear Variational Problems, Springer Series in Computational Physics, Springer-Verlag, New York, 1984. |
| [48] | V. Barbu, Nonlinear Differential Equations of Monotone Types in Banach Spaces, Springer Monographs in Mathematics, Springer, New York, 2010. |
| [49] | R. E. Showalter, Monotone Operators in Banach Space and Nonlinear Partial Differential Equations, American Mathematical Society, Providence, RI, 1997. |
| [50] |
C. Johnson, A convergence estimate for an approximation of a parabolic variational inequality, SIAM J. Numer. Anal., 13 (1976), 599–606. doi: 10.1137/0713050
|
| [51] | M. Ulbrich, Semismooth Newton Methods for Variational Inequalities and Constrained Optimization Problems in Function Spaces, Society for Industrial and Applied Mathematics (SIAM), Philadelphia, PA, 2011. |
| [52] | X. Chen, Convergence of numerical solutions to the Allen–Cahn equation, Appl. Anal., 69 (1998), 47–56. |
| [53] |
J. Shen, X. Yang, Numerical approximations of Allen–Cahn and Cahn–Hilliard equations, Discrete. Contin. Dyn. Syst., 28 (2010), 1669–1691. doi: 10.3934/dcds.2010.28.1669
|
| [54] | A. Visintin, Models of Phase Transitions, Birkhäuser, Birkhäuser Boston, 1996. |
| [55] | L. Tartar, Incompressible fluid flow in a porous medium–convergence of the homogenization process, in Nonhomogeneous Media and Vibration Theory, Springer-Verlag, Berlin, (1980), 368–377. |
| [56] | J. Bear, A. H. D. Cheng, Modeling Groundwater Flow and Contaminant Transport, Theory and Applications of Transport in Porous Media, Springer Netherlands, 2010. |
| [57] |
R. Schulz, N. Ray, S. Zech, A. Rupp, Peter Knabner, Beyond Kozeny–Carman: predicting the permeability in porous media, Transp. Porous Media, 130 (2019), 487–512. doi: 10.1007/s11242-019-01321-y
|
| [58] |
N. Ray, A. Rupp, R. Schulz, P. Knabner, Old and new approaches predicting the diffusion in porous media, Transp. Porous Media, 124 (2018), 803–824. doi: 10.1007/s11242-018-1099-x
|
| [59] | M. Peszynska, A. Trykozko, K. Augustson, Computational upscaling of inertia effects from porescale to mesoscale, in ICCS 2009 Proceedings, LNCS 5544, Part I (eds. G. Allen, J. Nabrzyski, E. Seidel, D. van Albada, J. Dongarra and P. Sloot), Springer-Verlag, Berlin-Heidelberg, (2009), 695–704. |
| [60] | M. Peszynska, A. Trykozko, W. Sobieski, Forchheimer law in computational and experimental studies of flow through porous media at porescale and mesoscale, GAKUTO Internat. Ser. Math. Sci. Appl., 32 (2010), 463–482. |
| [61] |
A. Trykozko, M. Peszynska, M. Dohnalik, Modeling non-Darcy flows in realistic porescale proppant geometries, Comput. Geotechnics, 71 (2016), 352–360. doi: 10.1016/j.compgeo.2015.08.011
|
| [62] | M. Peszynska, J. Umhoefer, C. Shin, Reduced model for properties of multiscale porous media with changing geometry, Computation, 9 (2021), 28. |
| [63] |
T. Arbogast, H. L. Lehr, Homogenization of a Darcy–Stokes system modeling vuggy porous media, Computat. Geosci., 10 (2006), 291–302. doi: 10.1007/s10596-006-9024-8
|
| [64] |
M. Krotkiewski, I. S. Ligaarden, K. A. Lie, D. W. Schmid, On the importance of the Stokes-Brinkman equations for computing effective permeability in karst reservoirs, Commun. Comput. Phys., 10 (2011), 1315–1332. doi: 10.4208/cicp.290610.020211a
|
| [65] | H. C. Brinkman, A calculation of the viscous force exerted by a flowing fluid on a dense swarm of particles, Appl. Sci. Res. Sec. A—Mechanics Heat Chem. Eng. Math. Methods, 1 (1947), 27-34. |
| [66] |
R. Guibert, P. Horgue, D. Gerald, M. Quintard, A comparison of various methods for the numerical evaluation of porous media permeability tensors from pore-scale geometry, Math. Geosci., 48 (2016), 329–347. doi: 10.1007/s11004-015-9587-9
|
| [67] |
J. Alberto Ochoa-Tapia, S. Whitaker, Momentum transfer at the boundary between a porous medium and a homogeneous fluid—i. theoretical development, Int. J. Heat Mass Transfer, 38 (1995), 2635–2646. doi: 10.1016/0017-9310(94)00346-W
|
| [68] |
J. Alberto Ochoa-Tapia, S. Whitaker, Momentum transfer at the boundary between a porous medium and a homogeneous fluid—ii. comparison with experiment, Int. J. Heat Mass Transfer, 38 (1995), 2647–2655. doi: 10.1016/0017-9310(94)00347-X
|
| [69] |
F. A. Morales, R. E. Showalter, A Darcy–Brinkman model of fractures in porous media, J. Math. Anal. Appl., 452 (2017), 1332–1358. doi: 10.1016/j.jmaa.2017.03.063
|
| [70] |
M. Discacciati, E. Miglio, A. Quarteroni, Mathematical and numerical models for coupling surface and groundwater flows, Appl. Numer. Math., 43 (2002), 57–74. doi: 10.1016/S0168-9274(02)00125-3
|
| [71] | A. Trykozko, M. Peszynska, Pore-scale simulations of pore clogging and upscaling with large velocities, GAKUTO Int. Ser., Math. Sci. Appl., 36 (2013), 277–300. |
| [72] |
W. Deng, M. Bayani Cardenas, M. F. Kirk, S. J. Altman, P. C. Bennett, Effect of permeable biofilm on micro- and macro-scale flow and transport in bioclogged pores, Environ. Sci. Technol., 47 (2013), 11092–11098. doi: 10.1021/es402596v
|
| [73] |
M. Thullner, J. Zeyer, W. Kinzelbach, Influence of microbial growth on hydraulic properties of pore networks, Transp. Porous Media, 49 (2002), 99–122. doi: 10.1023/A:1016030112089
|
| [74] | M. Peszynska, S. Sun, Reactive Transport Module TRCHEM in IPARS, Technical report, TICAM Report 01-32, 2001. |
| [75] | F. Saaf, A Study of Reactive Transport Phenomena in Porous Media, Ph.D thesis, Rice University, 1996. |
| [76] |
C. Ebmeyer, W. B. Liu, Finite element approximation of the fast diffusion and the porous medium equations, SIAM J. Numer. Anal., 46 (2008), 2393–2410. doi: 10.1137/060657728
|
| [77] |
M. Gokieli, N. Kenmochi, M. Niezgódka, Variational inequalities of Navier-Stokes type with time dependent constraints, J. Math. Anal. Appl., 449 (2017), 1229–1247. doi: 10.1016/j.jmaa.2016.12.048
|
| [78] |
M. Gokieli, N. Kenmochi, M. Niezgódka, Mathematical modeling of biofilm development, Nonlinear Anal. Real World Appl., 42 (2018), 422–447. doi: 10.1016/j.nonrwa.2018.01.005
|
| [79] | S. V. Patankar, Numerical Heat Transfer and Fluid Flow, Series in Computational Methods in Mechanics and Thermal Sciences, Routledge, 1980. |

Figures(19) / Tables(11)

Choah Shin, Azhar Alhammali, Lisa Bigler, Naren Vohra, Malgorzata Peszynska. Coupled flow and biomass-nutrient growth at pore-scale with permeable biofilm, adaptive singularity and multiple species[J]. Mathematical Biosciences and Engineering, 2021, 18(3): 2097-2149. doi: 10.3934/mbe.2021108







 DownLoad:
DownLoad: